
Mimivirus is a viral genus containing a single identified species named Acanthamoeba polyphaga mimivirus (APMV), or is a group of phylogenetically related large viruses (designated usually MimiN). In colloquial speech, APMV is more commonly referred to as just “mimivirus”. Until October 2011 it had largest capsid diameter of all known viruses, as well as a large and complex genome compared with other viruses.
Discovery
APMV was discovered serendipitously in 1992 within the amoeba Acanthamoeba polyphaga, after which it is named, during research into Legionellosis. The virus was observed in a gram stain and mistakenly thought to be a gram-positive bacterium. As a consequence it was named "Bradfordcoccus", after the district the amoeba was sourced from in Bradford, England. In 2003, researchers at the Université de la Méditerranée in Marseille, France published a paper in Science identifying the micro-organism as a virus.
Mimivirus may be a causative agent of some forms of pneumonia; this is based mainly on indirect evidence in the form of antibodies to the virus discovered in pneumonia patients. However, the classification of mimivirus as a pathogen is tenuous at present as there have been only a couple of papers published potentially linking mimivirus to actual cases of pneumonia. Indeed, a significant majority of pneumonia cases are due to unknown etiology.
Classification
Mimivirus has been placed into a viral family by the International Committee on Taxonomy of Viruses as a member of the Mimiviridae, and has been placed into Group I of the Baltimore classification system. Although not strictly a method of classification, Mimivirus joins a group of large viruses known as nucleocytoplasmic large DNA viruses (NCLDV). They are all large viruses which share both molecular characteristics and large genomes. The mimivirus genome also possesses 21 genes encoding homologs to proteins which are seen to be highly conserved in the majority of NCLDVs, and further work suggests that mimivirus is an early divergent of the general NCLDV group.
Structure
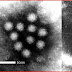
The mimivirus is larger than all previously discovered viruses, with a capsid diameter of 400 nm. Protein filaments measuring 100 nm project from the surface of the capsid, bringing the total length of the virus up to 600 nm. Variation in scientific literature renders these figures as highly approximate, with the "size" of the virion being casually listed as anywhere between 400 nm and 800 nm, depending on whether total length or capsid diameter is actually quoted. The capsid appears hexagonal under an electron microscope, therefore the capsid symmetry is icosahedral.[6] It does not appear to possess an outer viral envelope, suggesting that the virus does not exit the host cell by exocytosis.
The same team that discovered the mimivirus later discovered a slightly larger virus, dubbed the mamavirus, and the Sputnik virophage that infects it.
Mimivirus shares several morphological characteristics with all members of the NCLDV group of viruses. As an internal lipid layer surrounding the central core is present in all other NCLDV viruses, it has been suggested by M. Suzan-Monti et al. that this may also be present in mimivirus. The condensed central core of the virion appears as a dark region under the electron microscope. The large genome of the virus resides within this area.
Several mRNA transcripts can be recovered from purified virions. Like other NCLDVs, transcripts for DNA polymerase, a capsid protein and a TFII-like transcription factor were found. However, three distinct aminoacyl tRNA synthetase enzyme transcripts and four unknown mRNA molecules specific to mimivirus were also found. These pre-packaged transcripts can be translated without viral gene expression and are likely to be necessary to Mimivirus for replication. Other DNA viruses, such as the Human cytomegalovirus and Herpes simplex virus type-1, also feature pre-packaged mRNA transcripts.
Genome
The mimivirus genome is a linear, double-stranded molecule of DNA with 1,181,404 base pairs in length.[9] This makes it the largest viral genome in scientific knowledge, outstripping the next-largest virus genome of the Cafeteria roenbergensis virus by about 450,000 base pairs. In addition, it is larger than at least 30 cellular clades.
In addition to the large size of the genome, mimivirus possesses an estimated 979 protein-coding genes, far exceeding the minimum 4 genes required for viruses to exist (c.f. MS2 and Qβ viruses). Analysis of its genome revealed the presence of genes not seen in any other viruses, including aminoacyl tRNA synthetases, and other genes previously thought only to be encoded by cellular organisms. Like other large DNA viruses, mimivirus contains several genes for sugar, lipid and amino acid metabolism, as well as some metabolic genes not found in any other virus. Roughly 90% of the genome was of coding capacity, with the other 10% being “junk DNA”.
Replication
The stages of mimivirus replication are not well known, but as a minimum it is known that mimivirus attaches to a chemical receptor on the surface of an amoeba cell and is taken into the cell. Once inside, an eclipse phase begins, in which the virus disappears and all appears normal within the cell. After about four hours small accumulations can be seen in areas of the cell. Eight hours after infection many mimivirus virions are clearly visible within the cell. The cell cytoplasm continues to fill with newly synthesised virions and about 24 hours after initial infection the cell likely bursts open to release the new mimivirus.
virions.
Little is known about the details of this replication cycle, most obviously attachment to the cell surface and entry, viral core release, DNA replication, transcription, translation, assembly and release of progeny virions. However, scientists have established the general overview given above using electron micrographs of infected cells. These micrographs show mimivirus capsid assembly in the nucleus, acquisition of an inner lipid membrane via budding from the nucleus, and particles similar to those found in many other viruses, including all NCLDV members. These particles are known in other viruses as viral factories and allow efficient viral assembly by modifying large areas of the host cell.
Implications for defining "life"
Mimivirus possesses many characteristics which place it at the boundary of living and non-living. It is as large as several bacterial species, such as Rickettsia conorii and Tropheryma whipplei, possesses a genome of comparable size to several bacteria, including those above, and codes for products previously not thought to be encoded by viruses. In addition, mimivirus possesses genes coding for nucleotide and amino acid synthesis, which even some small obligate intracellular bacteria lack. This means that unlike these bacteria, mimivirus is not dependent on the host cell genome for coding the metabolic pathways for these products. They do however, lack genes for ribosomal proteins, making mimivirus dependent on a host cell for protein translation and energy metabolism. These factors combined have thrown scientists into debate over whether mimivirus is a distinct form of life, comparable on a domain scale to Eukarya, Archaea and Bacteria. Nevertheless, mimivirus does not exhibit the following characteristics, all of which are part of many conventional definitions of life: homeostasis, response to stimuli, growth in the normal sense of the term (instead replicating via self-assembly of individual components) or undergoing cellular division.
Because its lineage is very old and could have emerged prior to cellular organisms, mimivirus has added to the debate over the origins of life. Some genes unique to mimivirus, including those coding for the capsid, have been conserved in a variety of viruses which infect organisms from all domains - Eukarya, Archaea and Bacteria. This has been used to suggest that mimivirus is related to a type of DNA virus that emerged before cellular organisms and played a key role in the development of all life on Earth. An alternative
hypothesis is that there were three distinct types of DNA viruses that were involved in generating the three known domains of life.
References in popular culture
A "pseudo-mimi" drove much of the plot of Vernor Vinge's novel Rainbows End. In the book, it was a tailored virus used for highly effective and subtle mind control. The virus's relatively large genetic capacity was central to this idea.
Discovery
APMV was discovered serendipitously in 1992 within the amoeba Acanthamoeba polyphaga, after which it is named, during research into Legionellosis. The virus was observed in a gram stain and mistakenly thought to be a gram-positive bacterium. As a consequence it was named "Bradfordcoccus", after the district the amoeba was sourced from in Bradford, England. In 2003, researchers at the Université de la Méditerranée in Marseille, France published a paper in Science identifying the micro-organism as a virus.
Mimivirus may be a causative agent of some forms of pneumonia; this is based mainly on indirect evidence in the form of antibodies to the virus discovered in pneumonia patients. However, the classification of mimivirus as a pathogen is tenuous at present as there have been only a couple of papers published potentially linking mimivirus to actual cases of pneumonia. Indeed, a significant majority of pneumonia cases are due to unknown etiology.
Classification
Mimivirus has been placed into a viral family by the International Committee on Taxonomy of Viruses as a member of the Mimiviridae, and has been placed into Group I of the Baltimore classification system. Although not strictly a method of classification, Mimivirus joins a group of large viruses known as nucleocytoplasmic large DNA viruses (NCLDV). They are all large viruses which share both molecular characteristics and large genomes. The mimivirus genome also possesses 21 genes encoding homologs to proteins which are seen to be highly conserved in the majority of NCLDVs, and further work suggests that mimivirus is an early divergent of the general NCLDV group.
Structure
The mimivirus is larger than all previously discovered viruses, with a capsid diameter of 400 nm. Protein filaments measuring 100 nm project from the surface of the capsid, bringing the total length of the virus up to 600 nm. Variation in scientific literature renders these figures as highly approximate, with the "size" of the virion being casually listed as anywhere between 400 nm and 800 nm, depending on whether total length or capsid diameter is actually quoted. The capsid appears hexagonal under an electron microscope, therefore the capsid symmetry is icosahedral.[6] It does not appear to possess an outer viral envelope, suggesting that the virus does not exit the host cell by exocytosis.
The same team that discovered the mimivirus later discovered a slightly larger virus, dubbed the mamavirus, and the Sputnik virophage that infects it.
Mimivirus shares several morphological characteristics with all members of the NCLDV group of viruses. As an internal lipid layer surrounding the central core is present in all other NCLDV viruses, it has been suggested by M. Suzan-Monti et al. that this may also be present in mimivirus. The condensed central core of the virion appears as a dark region under the electron microscope. The large genome of the virus resides within this area.
Several mRNA transcripts can be recovered from purified virions. Like other NCLDVs, transcripts for DNA polymerase, a capsid protein and a TFII-like transcription factor were found. However, three distinct aminoacyl tRNA synthetase enzyme transcripts and four unknown mRNA molecules specific to mimivirus were also found. These pre-packaged transcripts can be translated without viral gene expression and are likely to be necessary to Mimivirus for replication. Other DNA viruses, such as the Human cytomegalovirus and Herpes simplex virus type-1, also feature pre-packaged mRNA transcripts.
Genome
The mimivirus genome is a linear, double-stranded molecule of DNA with 1,181,404 base pairs in length.[9] This makes it the largest viral genome in scientific knowledge, outstripping the next-largest virus genome of the Cafeteria roenbergensis virus by about 450,000 base pairs. In addition, it is larger than at least 30 cellular clades.
In addition to the large size of the genome, mimivirus possesses an estimated 979 protein-coding genes, far exceeding the minimum 4 genes required for viruses to exist (c.f. MS2 and Qβ viruses). Analysis of its genome revealed the presence of genes not seen in any other viruses, including aminoacyl tRNA synthetases, and other genes previously thought only to be encoded by cellular organisms. Like other large DNA viruses, mimivirus contains several genes for sugar, lipid and amino acid metabolism, as well as some metabolic genes not found in any other virus. Roughly 90% of the genome was of coding capacity, with the other 10% being “junk DNA”.
Replication
The stages of mimivirus replication are not well known, but as a minimum it is known that mimivirus attaches to a chemical receptor on the surface of an amoeba cell and is taken into the cell. Once inside, an eclipse phase begins, in which the virus disappears and all appears normal within the cell. After about four hours small accumulations can be seen in areas of the cell. Eight hours after infection many mimivirus virions are clearly visible within the cell. The cell cytoplasm continues to fill with newly synthesised virions and about 24 hours after initial infection the cell likely bursts open to release the new mimivirus.
virions.
Little is known about the details of this replication cycle, most obviously attachment to the cell surface and entry, viral core release, DNA replication, transcription, translation, assembly and release of progeny virions. However, scientists have established the general overview given above using electron micrographs of infected cells. These micrographs show mimivirus capsid assembly in the nucleus, acquisition of an inner lipid membrane via budding from the nucleus, and particles similar to those found in many other viruses, including all NCLDV members. These particles are known in other viruses as viral factories and allow efficient viral assembly by modifying large areas of the host cell.
Implications for defining "life"
Mimivirus possesses many characteristics which place it at the boundary of living and non-living. It is as large as several bacterial species, such as Rickettsia conorii and Tropheryma whipplei, possesses a genome of comparable size to several bacteria, including those above, and codes for products previously not thought to be encoded by viruses. In addition, mimivirus possesses genes coding for nucleotide and amino acid synthesis, which even some small obligate intracellular bacteria lack. This means that unlike these bacteria, mimivirus is not dependent on the host cell genome for coding the metabolic pathways for these products. They do however, lack genes for ribosomal proteins, making mimivirus dependent on a host cell for protein translation and energy metabolism. These factors combined have thrown scientists into debate over whether mimivirus is a distinct form of life, comparable on a domain scale to Eukarya, Archaea and Bacteria. Nevertheless, mimivirus does not exhibit the following characteristics, all of which are part of many conventional definitions of life: homeostasis, response to stimuli, growth in the normal sense of the term (instead replicating via self-assembly of individual components) or undergoing cellular division.
Because its lineage is very old and could have emerged prior to cellular organisms, mimivirus has added to the debate over the origins of life. Some genes unique to mimivirus, including those coding for the capsid, have been conserved in a variety of viruses which infect organisms from all domains - Eukarya, Archaea and Bacteria. This has been used to suggest that mimivirus is related to a type of DNA virus that emerged before cellular organisms and played a key role in the development of all life on Earth. An alternative
hypothesis is that there were three distinct types of DNA viruses that were involved in generating the three known domains of life.
References in popular culture
A "pseudo-mimi" drove much of the plot of Vernor Vinge's novel Rainbows End. In the book, it was a tailored virus used for highly effective and subtle mind control. The virus's relatively large genetic capacity was central to this idea.












0 komentar:
Posting Komentar